5.4 Experience Dependent Plasticity
Learning Objectives
By the end of this section, you should be able to
- 5.4.1 Describe and provide examples of experience-dependent plasticity during development.
- 5.4.2 Describe effects of environmental toxins and early experience during critical periods.
Early embryonic development provides the foundational unit of a brain with immature connections, but sensory cues can dramatically alter how brain maturation occurs. These sensory cues play a significant role in how neurons differentiate, how dendrites sprout, how neurons form and maintain synaptic connections, and how the brain produces final neural networks. At the early stages of development, our brain has a high degree of neural plasticity. Neural plasticity is the ability to adapt and reorganize in response to environmental stimuli. In fact, during certain periods of development called critical periods, environmental stimuli can lead to long-term positive or negative effects.
The brain's plasticity for refinement during critical periods can be either beneficial or detrimental. As an example of a beneficial experience, in rodent studies, enriched environments improve function of several brain regions when compared to animals homed in standard cages. These “enriched” living spaces provide settings that encourage exploration, cognitive engagement, social interaction, and physical activity in animals, enhancing plasticity. Rats raised in enriched environments show enhanced plasticity in several brain regions, even including sensory areas like the visual cortex, which shows an increase in connections and stronger circuits. Detrimental effects of negative environmental experiences are also, of course, possible. Monkeys and cats raised in restricted visual conditions, such as suturing an eye shut at a critical period of visual cortex growth or rearing in conditions without light, suffer permanent impairments in their visual abilities (Tian and Copenhagen, 2001; Vistamehr and Tian, 2004; Levelt and Hübener, 2012; Berry and Nedivi, 2016). These impairments include overcompensation in the plasticity to the eye receiving stimuli. Unfortunately, as we reach adulthood, the brain’s ability to reshape and grow new connections declines. At the same time, other factors can hasten the decline of cognitive plasticity, including environmental factors, hormones, and neurodegenerative diseases (Voss et al., 2017).
Embryonic and fetal period
Figure 5.29 defines critical periods in prenatal development. These periods are timepoints in prenatal development when specific organ systems of the embryo or fetus are particularly susceptible to environmental stimuli, meaning that negative stimuli can result in structural or functional brain malformations that impair function lifelong. CNS development begins embryonically at 3 weeks post-fertilization. The embryonic period lasts until 10 weeks, and encompasses neurulation, a critical period we have previously discussed as it relates to neural tube formation and closure. CNS development continues from 10 weeks to 38 weeks, which is defined as the period of the fetus.
The pioneering work of Charles Stockard and Hans Spemann has shown that there are stages that are particularly sensitive to environmental exposure and/or manipulation (Stockard, 1921; De Robertis, 2009). Stockard is known for his work with fish embryos, in which he found that introducing chemical agents at a specific stage would create what he called “monsters”. Figure 5.30 shows a drawing by Stockard of one of these fish after exposure to a teratogen during the critical embryonic period. This damage was permanent, and when replicated at other stages of development, there was essentially no similar defect seen, suggesting that there are specific prenatal stages that are very sensitive to teratogens. The reason for this susceptibility is that the fetus is experiencing rapid growth in size, weight, and muscle at early stages. Below we detail two common environmental exposures during fetal development that result in abnormal nervous system development, causing lifelong impairment in function.
Fetal alcohol syndrome
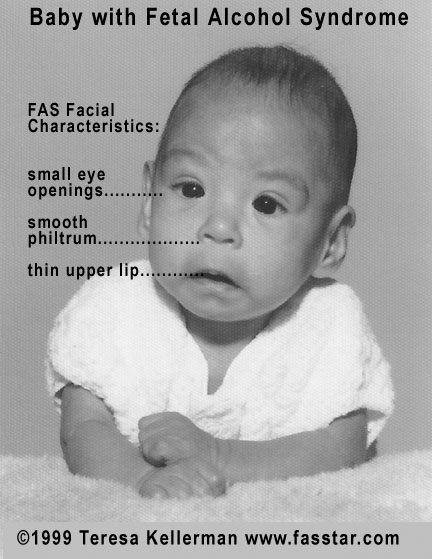
Exposure to alcohol during the fetal period can result in fetal alcohol syndrome (FAD) and fetal alcohol spectrum disorders (FASD). Alcohol is a teratogen, and its effects can lead to growth retardation, abnormal brain development, intellectual disability, and craniofacial abnormalities, which include a narrow forehead, flat midface, narrow eyelid openings, shortened nose, a long upper lip, and an absent philtrum (Figure 5.31; Sulik et al., 1981).
{kind=link}
FASD is the most common non-genetic form of intellectual disability, with broad symptoms, including poor cognition and behavioral phenotypes (Denny et al., 2017). The effects of FAD on growth and development can be observed in Figure 5.31. While the term “fetal” in FAD/FASD implies the effects of alcohol are focused only on the fetal period, alcohol can also have a negative impact earlier in pregnancy. When mice had a brief exposure to alcohol during embryonic development, they presented with severe facial malformations (Sulik et al., 1981). This period of embryonic development in mice is equivalent to approximately the third week of human gestation, a time point when many women aren’t aware of their pregnancy (Sulik et al., 1981). This is why it is recommended that individuals planning to become pregnant refrain from alcohol consumption.
Retinoic acid as a teratogen
Retinoic acid (RA) is a steroid hormone produced from vitamin A. Upon production, RA has several developmental responsibilities and can bind to RA receptors that together bind to DNA and change the genes that are expressed inside of a cell. Controlled levels of RA are essential for development, but excess intake of vitamin A or RA by pregnant women can lead to birth defects. These birth defects include microcephaly, spina bifida, hydrocephalus, and exencephaly. Importantly, drugs that contain forms of RA are frequently used to treat dermatological conditions including acne. Exposure to these drugs during pregnancy can predispose the fetus to CNS defects, as well as craniofacial abnormalities and shortened limbs (Berenguer et al., 2021).
2: Perinatal period
The perinatal period encompasses the time frame from childbirth up to 24 months. During this time, the CNS possesses tremendous plasticity, defined as the ability to remodel or reorganize based on experience. Such plasticity is very robust from birth to 3 years and gives rise to developmental critical periods. While the early brain maintains a high degree of plasticity, it is important to note that plasticity is maintained into adulthood, although it does decrease as we age. As discussed previously, significant synapse refinement occurs in the form of loss of poly-innervation and pruning during this period of development. Below, we will discuss several examples of how long-term brain function can be shaped by experiences during this perinatal period.
Perinatal brain development and parental care
One of the most tragic consequences of the perinatal critical period for CNS development is the long-term and severe impact that neglect or abuse during this time can have. Figure 5.32 highlights some of the major brain regions where disrupted growth is evident years after childhood abuse/neglect. They include regions important for emotion (amygdala), memory (hippocampus), and executive function (prefrontal cortex). These kinds of extreme examples show how experience in early life can disrupt the structure of the brain long-term.
Neuroscience in the lab: Uncovering mechanisms of parental care effects on brain development
The importance of perinatal care experiences on brain development cannot be underemphasized. But how do these nurturing events (or their absence) in infancy and childhood affect our adult behavior? Studies in rats have provided some insight into how early life care can affect adult behaviors by shaping the stress response. Maternal rats provide nurturing to their offspring through licking and grooming. Rat moms are not all equal and some provide more licking and grooming than others, even in the controlled conditions of the lab. Meaney and Szyf (2022) took advantage of this natural variation to study the consequences of maternal behavior on the long-term development of their pups. Remarkably, they found that adult rats that received more grooming attention as pups responded better to stressful events later in life (Liu et al., 1997) (see Chapter 12 Stress). Specifically, they secreted less stress hormones in response to a stressor created by the researchers. The key to their reduced hormonal response was a long-lasting increase in stress hormone receptors. A high number of these receptors inhibits the excessive production of stress hormones leading to reduced hormonal stress response. This example shows one mechanism by which perinatal experience can lead to life-long changes that impact brain function, in this case the response to stress.
Perinatal critical period for language development
Language development is particularly sensitive to input during the perinatal and juvenile time period. An infamous story of neglect, given the pseudonym “Genie”, has helped further understand the importance of language development during critical periods (Salus and Curtiss, 1979; Rymer, 1993). The unfortunate events this child endured were extreme deprivation of sensory experience. Genie was confined to a small room, isolated, and tied to a potty seat, every day up to 13 years of age. The child was severely deprived from receiving any sensory stimuli, to the point where her father prohibited that anyone talk to Genie, instead only barking or growling at her was allowed. For many years after she was found, Genie received help among many therapists to aid in her development. Through extensive therapy, she was able to learn basic words and word combinations, but she still lacked the ability to speak in full sentences or understand the concept of assigning different words to the same meaning.
Genie’s story is anecdotal evidence as to the importance of critical periods during language development and is somewhat complicated by the severity of her deficits in other aspects of cognitive function. More definitive evidence indicating the importance of a critical period for language development has been established through the study of the birdsong. Oscine songbirds use the birdsong to attract their mates. In order to develop the appropriate song, juvenile birds must be exposed to their song by an adult male, memorize the song, and then recapitulate it. While birdsong is not the same as human language, it has some similarities as a form of vocal learning. If a bird is not exposed to the correct song within the first 2 months of hatching or exposed to a different song during this critical period, the bird will not develop the appropriate song and can fail to attract a mate (Doupe and Kuhl, 1999).
Oscine birds and humans share the need for environmental exposure for proper language development. Children develop proper syntax within the first year of life (Friedmann and Rusou, 2015) and it is well-established that learning a language happens spontaneously among children, but not in adults. Adults mostly must explicitly study a new language to learn it and even then, grammar, syntax and pronunciation are typically not as proficient as those who learned before adolescence. This early childhood critical period for language acquisition likely relies on the higher synaptic plasticity in language centers of the brain during this phase of development, when synapses are being established and refined.
Perinatal critical period in visual development
Perinatal sensory experience is required for the development of the visual system at several levels. One prominent example of this is development of ocular dominance in the primary visual cortex (called V1 for short). At birth, the layers of the primary visual cortex are innervated by axons from both eyes. However, by adulthood through visual stimulation, the innervations are remodeled and segregated between layers to produce ocular dominance columns (Stryker and Harris, 1986; Levelt and Hübener, 2012; Berry and Nedivi, 2016). Within an ocular dominance column, the cells respond preferentially to light being shown in one eye over the other. This process of refinement to create preference for input from one eye requires visual input, as was first demonstrated by studies in cats.
David Hubel and Torsten Wiesel performed the first key studies to establish critical periods of vision development (Wiesel and Hubel, 1963; Hubel and Wiesel, 1970; Levelt and Hübener, 2012; Berry and Nedivi, 2016). To study how exposure to light influences formation of cortical cell preference for responding to input from one eye or the other, Hubel and Wiesel used the simple model of suturing shut a single eye of feline subjects at different times. They measured ocular preference of cortical cells by recording from individual cells in the cat visual cortex while shining a light on one eye or the other. Figure 5.33 describes their findings.
In an adult cat that never had an eye sutured shut (i.e., a normal cat), they found a mixture of visual cortical cells that responded preferentially to the right eye, left eye, or a bit of both. However, if a cat experienced just 3 to 4 days of visual deprivation when they were a kitten (perinatally), almost all their visual cortical cells responded only to light from the unsutured eye (Hubel and Wiesel, 1970). The formerly sutured eye was effectively blind because it did not stimulate any cortical cells. Importantly, if the eye was sutured shut during adulthood, there was no effect (Figure 5.33), effectively confirming a critical period for vision development. The first few weeks of life are a critical period for this part of visual development because this is when inputs to the visual cortex are forming synapses on V1 cortical cells. Cortical cells end up only connected to inputs deriving from the open eye because of a lack of competition from the axons in the sutured eye. Opening the eye later can’t fix that as the critical period for synapse formation has passed.
The significance of visual stimulation and critical periods has been further substantiated by a famous case study in Oliver Sacks’ renowned publication (1985). There he described a human patient by the surname Fergal, who was born blind and received surgery to regain vision after 50 years without it. Despite the success of regaining vision, Fergal lacked the early experiences necessary for synaptic reorganization and proper vision development (Sacks, 1985). This resulted in an influx of stimuli, as his other senses were heightened. As Sacks (1985) describes, “There were no visual memories to support a perception; there was no world of experience and meaning awaiting him. He saw, but what he saw had no coherence. His retina and optic nerve were active, transmitting impulses, but his brain could make no sense of them.” A second example of sensory critical periods and intervention includes the usage of cochlear implants (see Chapter 7 Hearing and Balance). The efficacy of cochlear implants is widely affected by the age of the patient. For example, the efficacy associated with cochlear implants in children at a young age, with shorter periods of auditory deprivation, is much higher than those with prolonged auditory deprivation (Pisoni et al. 1999).
Plasticity during adolescence
Adolescence refers to the stage of development in between childhood and adulthood. Thus far, we have discussed robust examples of plasticity during infancy and early development. However, the adolescent brain exhibits tremendous neural development, which makes it highly plastic and susceptible to environmental influence. Adolescence is particularly characterized by refinement in connectivity between major brain regions. For example, the connections between the frontal lobe and the amygdala are subject to extensive experience dependent refinement during adolescence. The prefrontal cortex (PFC) is responsible for higher order function and decision making called executive function and the amygdala regulates fear, aggression, and emotion. Adolescent decision making is also influenced by the immaturity of the reward centers in the brain, which include the nucleus accumbens (Spear, 2000), which contains dopaminergic input. The relative immaturity of these 3 brain regions during adolescence leads to more impulsive, reward based, and risk like behaviors relative to adults (Ernst, 2014). It also underlies their enhanced vulnerability/plasticity during adolescence. Studies have shown that chronic stress and psychological distress can have brain region specific impacts during adolescence.For example, abnormal development of the PFC can affect working memory, cognitive flexibility, and self-control.
Adolescence is also a unique critical period for the effects of substance misuse on specific regions of the brain. For instance, substance misuse that occurs during early adolescence, around ages 14-16, is associated with reduction in the volume of frontal cortex gray matter, whereas misuse in childhood selectively impacts the hippocampal gray matter volume (Larsen & Luna, 2018). Such differences can be explained by the different time points in development where growth is more prevalent for each of these two brain regions.